.
Bands result from membrane strip incubation.")
| 製品: | hnRNP U Antibody |
| カタログ: | DF13346 |
| タンパク質の説明: | Rabbit polyclonal antibody to hnRNP U |
| アプリケーション: | WB |
| 反応性: | Human, Mouse, Rat |
| 予測: | Pig, Bovine, Horse, Sheep, Rabbit, Dog, Chicken, Xenopus |
| 分子量: | 90kDa(Observed); 91kD(Calculated). |
| ユニプロット: | Q00839 |
| RRID: | AB_2846365 |
Control Products
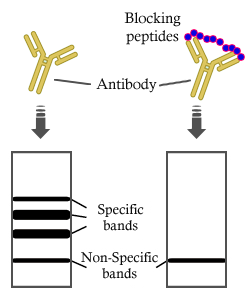
製品説明
*The optimal dilutions should be determined by the end user. For optimal experimental results, antibody reuse is not recommended.
*Tips:
WB: For western blot detection of denatured protein samples. IHC: For immunohistochemical detection of paraffin sections (IHC-p) or frozen sections (IHC-f) of tissue samples. IF/ICC: For immunofluorescence detection of cell samples. ELISA(peptide): For ELISA detection of antigenic peptide.
引用形式: Affinity Biosciences Cat# DF13346, RRID:AB_2846365.
折りたたみ/展開
Heterogeneous nuclear ribonucleoprotein U; hnRNP U; hnRNP U protein; HNRNPU; hnRNPU protein; HNRPU; HNRPU_HUMAN; p120; p120 nuclear protein; pp120; SAF A; SAF-A; SAFA; Scaffold attachment factor A; U21.1;
免疫原
A synthesized peptide derived from human hnRNP U, corresponding to a region within C-terminal amino acids.
- Q00839 HNRPU_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MSSSPVNVKKLKVSELKEELKKRRLSDKGLKAELMERLQAALDDEEAGGRPAMEPGNGSLDLGGDSAGRSGAGLEQEAAAGGDEEEEEEEEEEEGISALDGDQMELGEENGAAGAADSGPMEEEEAASEDENGDDQGFQEGEDELGDEEEGAGDENGHGEQQPQPPATQQQQPQQQRGAAKEAAGKSSGPTSLFAVTVAPPGARQGQQQAGGKKKAEGGGGGGRPGAPAAGDGKTEQKGGDKKRGVKRPREDHGRGYFEYIEENKYSRAKSPQPPVEEEDEHFDDTVVCLDTYNCDLHFKISRDRLSASSLTMESFAFLWAGGRASYGVSKGKVCFEMKVTEKIPVRHLYTKDIDIHEVRIGWSLTTSGMLLGEEEFSYGYSLKGIKTCNCETEDYGEKFDENDVITCFANFESDEVELSYAKNGQDLGVAFKISKEVLAGRPLFPHVLCHNCAVEFNFGQKEKPYFPIPEEYTFIQNVPLEDRVRGPKGPEEKKDCEVVMMIGLPGAGKTTWVTKHAAENPGKYNILGTNTIMDKMMVAGFKKQMADTGKLNTLLQRAPQCLGKFIEIAARKKRNFILDQTNVSAAAQRRKMCLFAGFQRKAVVVCPKDEDYKQRTQKKAEVEGKDLPEHAVLKMKGNFTLPEVAECFDEITYVELQKEEAQKLLEQYKEESKKALPPEKKQNTGSKKSNKNKSGKNQFNRGGGHRGRGGFNMRGGNFRGGAPGNRGGYNRRGNMPQRGGGGGGSGGIGYPYPRAPVFPGRGSYSNRGNYNRGGMPNRGNYNQNFRGRGNNRGYKNQSQGYNQWQQGQFWGQKPWSQHYHQGYY
種類予測
Score>80(red) has high confidence and is suggested to be used for WB detection. *The prediction model is mainly based on the alignment of immunogen sequences, the results are for reference only, not as the basis of quality assurance.
High(score>80) Medium(80>score>50) Low(score<50) No confidence
研究背景
DNA- and RNA-binding protein involved in several cellular processes such as nuclear chromatin organization, telomere-length regulation, transcription, mRNA alternative splicing and stability, Xist-mediated transcriptional silencing and mitotic cell progression. Plays a role in the regulation of interphase large-scale gene-rich chromatin organization through chromatin-associated RNAs (caRNAs) in a transcription-dependent manner, and thereby maintains genomic stability. Required for the localization of the long non-coding Xist RNA on the inactive chromosome X (Xi) and the subsequent initiation and maintenance of X-linked transcriptional gene silencing during X-inactivation (By similarity). Plays a role as a RNA polymerase II (Pol II) holoenzyme transcription regulator. Promotes transcription initiation by direct association with the core-TFIIH basal transcription factor complex for the assembly of a functional pre-initiation complex with Pol II in a actin-dependent manner. Blocks Pol II transcription elongation activity by inhibiting the C-terminal domain (CTD) phosphorylation of Pol II and dissociates from Pol II pre-initiation complex prior to productive transcription elongation. Positively regulates CBX5-induced transcriptional gene silencing and retention of CBX5 in the nucleus. Negatively regulates glucocorticoid-mediated transcriptional activation. Key regulator of transcription initiation and elongation in embryonic stem cells upon leukemia inhibitory factor (LIF) signaling (By similarity). Involved in the long non-coding RNA H19-mediated Pol II transcriptional repression. Participates in the circadian regulation of the core clock component ARNTL/BMAL1 transcription (By similarity). Plays a role in the regulation of telomere length. Plays a role as a global pre-mRNA alternative splicing modulator by regulating U2 small nuclear ribonucleoprotein (snRNP) biogenesis. Plays a role in mRNA stability. Component of the CRD-mediated complex that promotes MYC mRNA stabilization. Enhances the expression of specific genes, such as tumor necrosis factor TNFA, by regulating mRNA stability, possibly through binding to the 3'-untranslated region (UTR). Plays a role in mitotic cell cycle regulation. Involved in the formation of stable mitotic spindle microtubules (MTs) attachment to kinetochore, spindle organization and chromosome congression. Phosphorylation at Ser-59 by PLK1 is required for chromosome alignement and segregation and progression through mitosis. Contributes also to the targeting of AURKA to mitotic spindle MTs. Binds to double- and single-stranded DNA and RNA, poly(A), poly(C) and poly(G) oligoribonucleotides. Binds to chromatin-associated RNAs (caRNAs). Associates with chromatin to scaffold/matrix attachment region (S/MAR) elements in a chromatin-associated RNAs (caRNAs)-dependent manner. Binds to the Xist RNA. Binds the long non-coding H19 RNA. Binds to SMN1/2 pre-mRNAs at G/U-rich regions. Binds to small nuclear RNAs (snRNAs). Binds to the 3'-UTR of TNFA mRNA. Binds (via RNA-binding RGG-box region) to the long non-coding Xist RNA; this binding is direct and bridges the Xist RNA and the inactive chromosome X (Xi) (By similarity). Also negatively regulates embryonic stem cell differentiation upon LIF signaling (By similarity). Required for embryonic development (By similarity). Binds to brown fat long non-coding RNA 1 (Blnc1); facilitates the recruitment of Blnc1 by ZBTB7B required to drive brown and beige fat development and thermogenesis (By similarity).
(Microbial infection) Negatively regulates immunodeficiency virus type 1 (HIV-1) replication by preventing the accumulation of viral mRNA transcripts in the cytoplasm.
Cleaved at Asp-100 by CASP3 during T-cell apoptosis, resulting in a loss of DNA- and chromatin-binding activities.
Extensively phosphorylated. Phosphorylated on Ser-59 by PLK1 and dephosphorylated by protein phosphatase 2A (PP2A) in mitosis.
Arg-739 is dimethylated, probably to asymmetric dimethylarginine (Ref.8). Arg-733 is dimethylated, probably to asymmetric dimethylarginine (By similarity).
Citrullinated by PADI4.
Nucleus. Nucleus matrix. Chromosome. Nucleus speckle. Cytoplasm>Cytoskeleton>Microtubule organizing center>Centrosome. Chromosome>Centromere>Kinetochore. Cytoplasm>Cytoskeleton>Spindle. Cytoplasm>Cytoskeleton>Spindle pole. Midbody. Cytoplasm. Cell surface. Cytoplasmic granule.
Note: Localizes at inactive X chromosome (Xi) regions (PubMed:11003645, PubMed:14608463, PubMed:15563465). Localizes in the nucleus during interphase (PubMed:21242313). At metaphase, localizes with mitotic spindle microtubules (MTs) (PubMed:21242313). At anaphase, localizes in the mitotic spindle midzone (PubMed:21242313). Localizes in spindle MTs proximal to spindle poles in a TPX2- and AURKA-dependent manner (PubMed:21242313). The Ser-59 phosphorylated form localizes to centrosomes during prophase and metaphase, to mitotic spindles in anaphase and to the midbody during cytokinesis (PubMed:25986610). Colocalizes with SMARCA4 in the nucleus (By similarity). Colocalizes with CBX5 in the nucleus (PubMed:19617346). Colocalizes with NR3C1 in nuclear speckles (PubMed:9353307). Localized in cytoplasmic ribonucleoprotein (RNP) granules containing untranslated mRNAs (PubMed:17289661).
Widely expressed.
The SAP domain is necessary for specific binding to nuclear scaffold/matrix attachment region (S/MAR) elements in DNA (PubMed:9405365, PubMed:11003645). The RNA-binding RGG-box region is necessary for its association with inactive X chromosome (Xi) regions and to chromatin-associated RNAs (caRNAs) (PubMed:14608463, PubMed:28622508). Both the DNA-binding domain SAP and the RNA-binding RGG-box region are necessary for the localization of Xist RNA on the Xi (By similarity). The ATPase and RNA-binding RGG-box regions are necessary for oligomerization (PubMed:28622508).
研究領域
· Genetic Information Processing > Transcription > Spliceosome.
Restrictive clause
Affinity Biosciences tests all products strictly. Citations are provided as a resource for additional applications that have not been validated by Affinity Biosciences. Please choose the appropriate format for each application and consult Materials and Methods sections for additional details about the use of any product in these publications.
For Research Use Only.
Not for use in diagnostic or therapeutic procedures. Not for resale. Not for distribution without written consent. Affinity Biosciences will not be held responsible for patent infringement or other violations that may occur with the use of our products. Affinity Biosciences, Affinity Biosciences Logo and all other trademarks are the property of Affinity Biosciences LTD.