, using Tristetraprolin Antibody at 1/1000 dilution.
5ug/NC membrane strip.
Exposure for 5s with Affinity™ ECL Kit(#KF8003).
Bands result from membrane strip incubation.")



Control Products
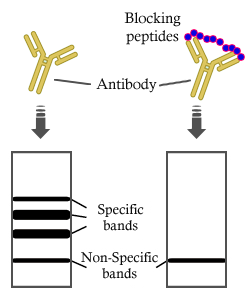
製品説明
*The optimal dilutions should be determined by the end user. For optimal experimental results, antibody reuse is not recommended.
*Tips:
WB: For western blot detection of denatured protein samples. IHC: For immunohistochemical detection of paraffin sections (IHC-p) or frozen sections (IHC-f) of tissue samples. IF/ICC: For immunofluorescence detection of cell samples. ELISA(peptide): For ELISA detection of antigenic peptide.
引用形式: Affinity Biosciences Cat# DF13378, RRID:AB_2846397.
折りたたみ/展開
G0/G1 switch regulatory protein 24; G0S24; GOS24; Growth factor-inducible nuclear protein NUP475; NUP475; Protein TIS11A; RNF162A; TIS 11; TIS11; TIS11A; Tristetraprolin; Tristetraproline; TTP; TTP_HUMAN; Zfp-36; ZFP36; Zinc finger protein 36; Zinc finger protein 36 homolog; Zinc finger protein 36, C3H type, homolog (mouse); Zinc finger protein, C3H type, 36 homolog;
免疫原
A synthesized peptide derived from human Tristetraprolin, corresponding to a region within the internal amino acids.
Expressed in both basal and suprabasal epidermal layers (PubMed:27182009). Expressed in epidermal keratinocytes (PubMed:27182009). Expressed strongly in mature dendritic cells (PubMed:18367721). Expressed in immature dendritic cells (at protein level) (PubMed:18367721).
- P26651 TTP_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MDLTAIYESLLSLSPDVPVPSDHGGTESSPGWGSSGPWSLSPSDSSPSGVTSRLPGRSTSLVEGRSCGWVPPPPGFAPLAPRLGPELSPSPTSPTATSTTPSRYKTELCRTFSESGRCRYGAKCQFAHGLGELRQANRHPKYKTELCHKFYLQGRCPYGSRCHFIHNPSEDLAAPGHPPVLRQSISFSGLPSGRRTSPPPPGLAGPSLSSSSFSPSSSPPPPGDLPLSPSAFSAAPGTPLARRDPTPVCCPSCRRATPISVWGPLGGLVRTPSVQSLGSDPDEYASSGSSLGGSDSPVFEAGVFAPPQPVAAPRRLPIFNRISVSE
研究背景
Zinc-finger RNA-binding protein that destabilizes several cytoplasmic AU-rich element (ARE)-containing mRNA transcripts by promoting their poly(A) tail removal or deadenylation, and hence provide a mechanism for attenuating protein synthesis. Acts as an 3'-untranslated region (UTR) ARE mRNA-binding adapter protein to communicate signaling events to the mRNA decay machinery. Recruits deadenylase CNOT7 (and probably the CCR4-NOT complex) via association with CNOT1, and hence promotes ARE-mediated mRNA deadenylation. Functions also by recruiting components of the cytoplasmic RNA decay machinery to the bound ARE-containing mRNAs. Self regulates by destabilizing its own mRNA. Binds to 3'-UTR ARE of numerous mRNAs and of its own mRNA. Plays a role in anti-inflammatory responses; suppresses tumor necrosis factor (TNF)-alpha production by stimulating ARE-mediated TNF-alpha mRNA decay and several other inflammatory ARE-containing mRNAs in interferon (IFN)- and/or lipopolysaccharide (LPS)-induced macrophages (By similarity). Plays also a role in the regulation of dendritic cell maturation at the post-transcriptional level, and hence operates as part of a negative feedback loop to limit the inflammatory response. Promotes ARE-mediated mRNA decay of hypoxia-inducible factor HIF1A mRNA during the response of endothelial cells to hypoxia. Positively regulates early adipogenesis of preadipocytes by promoting ARE-mediated mRNA decay of immediate early genes (IEGs) (By similarity). Negatively regulates hematopoietic/erythroid cell differentiation by promoting ARE-mediated mRNA decay of the transcription factor STAT5B mRNA. Plays a role in maintaining skeletal muscle satellite cell quiescence by promoting ARE-mediated mRNA decay of the myogenic determination factor MYOD1 mRNA (By similarity). Associates also with and regulates the expression of non-ARE-containing target mRNAs at the post-transcriptional level, such as MHC class I mRNAs. Participates in association with argonaute RISC catalytic components in the ARE-mediated mRNA decay mechanism; assists microRNA (miRNA) targeting ARE-containing mRNAs. May also play a role in the regulation of cytoplasmic mRNA decapping; enhances decapping of ARE-containing RNAs, in vitro. Involved in the delivery of target ARE-mRNAs to processing bodies (PBs). In addition to its cytosolic mRNA-decay function, affects nuclear pre-mRNA processing (By similarity). Negatively regulates nuclear poly(A)-binding protein PABPN1-stimulated polyadenylation activity on ARE-containing pre-mRNA during LPS-stimulated macrophages (By similarity). Also involved in the regulation of stress granule (SG) and P-body (PB) formation and fusion (By similarity). Plays a role in the regulation of keratinocyte proliferation, differentiation and apoptosis. Plays a role as a tumor suppressor by inhibiting cell proliferation in breast cancer cells.
(Microbial infection) Negatively regulates HTLV-1 TAX-dependent transactivation of viral long terminal repeat (LTR) promoter.
Phosphorylated. Phosphorylation at serine and/or threonine residues occurs in a p38 MAPK- and MAPKAPK2-dependent manner. Phosphorylated by MAPKAPK2 at Ser-60 and Ser-186; phosphorylation increases its stability and cytoplasmic localization, promotes binding to 14-3-3 adapter proteins and inhibits the recruitment of cytoplasmic CCR4-NOT and PAN2-PAN3 deadenylase complexes to the mRNA decay machinery, thereby inhibiting ZFP36-induced ARE-containing mRNA deadenylation and decay processes. Phosphorylation by MAPKAPK2 does not impair ARE-containing RNA-binding. Phosphorylated in a MAPKAPK2- and p38 MAPK-dependent manner upon skeletal muscle satellite cell activation; this phosphorylation inhibits ZFP36-mediated mRNA decay activity, and hence stabilizes MYOD1 mRNA (By similarity). Phosphorylated by MAPK1 upon mitogen stimulation (By similarity). Phosphorylated at Ser-66 and Ser-93; these phosphorylations increase in a SH3KBP1-dependent manner. Phosphorylated at serine and threonine residues in a pyruvate kinase PKM- and p38 MAPK-dependent manner. Phosphorylation at Ser-60 may participate in the PKM-mediated degradation of ZFP36 in a p38 MAPK-dependent manner. Dephosphorylated by serine/threonine phosphatase 2A at Ser-186 (By similarity).
Ubiquitinated; pyruvate kinase (PKM)-dependent ubiquitination leads to proteasomal degradation through a p38 MAPK signaling pathway.
Nucleus. Cytoplasm. Cytoplasmic granule. Cytoplasm>P-body.
Note: Shuttles between nucleus and cytoplasm in a CRM1-dependent manner (By similarity). Localized predominantly in the cytoplasm in a p38 MAPK- and YWHAB-dependent manner (By similarity). Colocalizes with SH3KBP1 and MAP3K4 in the cytoplasm (PubMed:20221403). Component of cytoplasmic stress granules (SGs) (By similarity). Localizes to cytoplasmic stress granules upon energy starvation (PubMed:15014438). Localizes in processing bodies (PBs) (PubMed:17369404). Excluded from stress granules in a phosphorylation MAPKAPK2-dependent manner (By similarity). Shuttles in and out of both cytoplasmic P-body and SGs (By similarity).
Nucleus. Cytoplasm.
Note: (Microbial infection) Colocalizes with HTLV-1 TAX in the nucleus and the cytoplasm in a region surrounding the nucleus.
Expressed in both basal and suprabasal epidermal layers. Expressed in epidermal keratinocytes. Expressed strongly in mature dendritic cells. Expressed in immature dendritic cells (at protein level).
The C3H1-type zinc finger domains are necessary for ARE-binding activity (PubMed:10330172).
研究領域
· Human Diseases > Infectious diseases: Viral > HTLV-I infection.
Restrictive clause
Affinity Biosciences tests all products strictly. Citations are provided as a resource for additional applications that have not been validated by Affinity Biosciences. Please choose the appropriate format for each application and consult Materials and Methods sections for additional details about the use of any product in these publications.
For Research Use Only.
Not for use in diagnostic or therapeutic procedures. Not for resale. Not for distribution without written consent. Affinity Biosciences will not be held responsible for patent infringement or other violations that may occur with the use of our products. Affinity Biosciences, Affinity Biosciences Logo and all other trademarks are the property of Affinity Biosciences LTD.