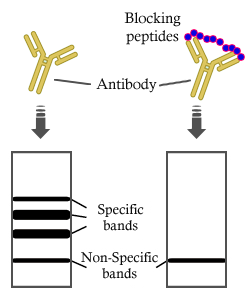
, using PML Antibody. The lane on the left was treated with blocking peptide.")
.
Bands result from membrane strip incubation.")
, using PML Antibody at 1/1000 dilution.
5ug/NC membrane strip.
Exposure for 10s with Affinity™ ECL Kit(#KF8001).
Bands result from membrane strip incubation.")
, using PML Antibody at 1/1000 dilution.
5ug/NC membrane strip.
Exposure for 20s with Affinity™ ECL Kit(#KF8001).
Bands result from membrane strip incubation.")






製品説明
*The optimal dilutions should be determined by the end user. For optimal experimental results, antibody reuse is not recommended.
*Tips:
WB: For western blot detection of denatured protein samples. IHC: For immunohistochemical detection of paraffin sections (IHC-p) or frozen sections (IHC-f) of tissue samples. IF/ICC: For immunofluorescence detection of cell samples. ELISA(peptide): For ELISA detection of antigenic peptide.
引用形式: Affinity Biosciences Cat# DF6318, RRID:AB_2838284.
折りたたみ/展開
Acure promyelocytic leukemia, inducer of; MYL; Pml; PML_HUMAN; PP8675; Probable transcription factor PML; Promyelocytic leukemia; Promyelocytic leukemia inducer of; Promyelocytic leukemia protein; Protein PML; RING finger protein 71; RNF 71; RNF71; TRIM 19; Tripartite motif protein TRIM19; Tripartite motif-containing protein 19;
免疫原
A synthesized peptide derived from human PML, corresponding to a region within N-terminal amino acids.
- P29590 PML_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MEPAPARSPRPQQDPARPQEPTMPPPETPSEGRQPSPSPSPTERAPASEEEFQFLRCQQCQAEAKCPKLLPCLHTLCSGCLEASGMQCPICQAPWPLGADTPALDNVFFESLQRRLSVYRQIVDAQAVCTRCKESADFWCFECEQLLCAKCFEAHQWFLKHEARPLAELRNQSVREFLDGTRKTNNIFCSNPNHRTPTLTSIYCRGCSKPLCCSCALLDSSHSELKCDISAEIQQRQEELDAMTQALQEQDSAFGAVHAQMHAAVGQLGRARAETEELIRERVRQVVAHVRAQERELLEAVDARYQRDYEEMASRLGRLDAVLQRIRTGSALVQRMKCYASDQEVLDMHGFLRQALCRLRQEEPQSLQAAVRTDGFDEFKVRLQDLSSCITQGKDAAVSKKASPEAASTPRDPIDVDLPEEAERVKAQVQALGLAEAQPMAVVQSVPGAHPVPVYAFSIKGPSYGEDVSNTTTAQKRKCSQTQCPRKVIKMESEEGKEARLARSSPEQPRPSTSKAVSPPHLDGPPSPRSPVIGSEVFLPNSNHVASGAGEAEERVVVISSSEDSDAENSSSRELDDSSSESSDLQLEGPSTLRVLDENLADPQAEDRPLVFFDLKIDNETQKISQLAAVNRESKFRVVIQPEAFFSIYSKAVSLEVGLQHFLSFLSSMRRPILACYKLWGPGLPNFFRALEDINRLWEFQEAISGFLAALPLIRERVPGASSFKLKNLAQTYLARNMSERSAMAAVLAMRDLCRLLEVSPGPQLAQHVYPFSSLQCFASLQPLVQAAVLPRAEARLLALHNVSFMELLSAHRRDRQGGLKKYSRYLSLQTTTLPPAQPAFNLQALGTYFEGLLEGPALARAEGVSTPLAGRGLAERASQQS
研究背景
Functions via its association with PML-nuclear bodies (PML-NBs) in a wide range of important cellular processes, including tumor suppression, transcriptional regulation, apoptosis, senescence, DNA damage response, and viral defense mechanisms. Acts as the scaffold of PML-NBs allowing other proteins to shuttle in and out, a process which is regulated by SUMO-mediated modifications and interactions. Isoform PML-4 has a multifaceted role in the regulation of apoptosis and growth suppression: activates RB1 and inhibits AKT1 via interactions with PP1 and PP2A phosphatases respectively, negatively affects the PI3K pathway by inhibiting MTOR and activating PTEN, and positively regulates p53/TP53 by acting at different levels (by promoting its acetylation and phosphorylation and by inhibiting its MDM2-dependent degradation). Isoform PML-4 also: acts as a transcriptional repressor of TBX2 during cellular senescence and the repression is dependent on a functional RBL2/E2F4 repressor complex, regulates double-strand break repair in gamma-irradiation-induced DNA damage responses via its interaction with WRN, acts as a negative regulator of telomerase by interacting with TERT, and regulates PER2 nuclear localization and circadian function. Isoform PML-6 inhibits specifically the activity of the tetrameric form of PKM. The nuclear isoforms (isoform PML-1, isoform PML-2, isoform PML-3, isoform PML-4 and isoform PML-5) in concert with SATB1 are involved in local chromatin-loop remodeling and gene expression regulation at the MHC-I locus. Isoform PML-2 is required for efficient IFN-gamma induced MHC II gene transcription via regulation of CIITA. Cytoplasmic PML is involved in the regulation of the TGF-beta signaling pathway. PML also regulates transcription activity of ELF4 and can act as an important mediator for TNF-alpha- and IFN-alpha-mediated inhibition of endothelial cell network formation and migration.
Exhibits antiviral activity against both DNA and RNA viruses. The antiviral activity can involve one or several isoform(s) and can be enhanced by the permanent PML-NB-associated protein DAXX or by the recruitment of p53/TP53 within these structures. Isoform PML-4 restricts varicella zoster virus (VZV) via sequestration of virion capsids in PML-NBs thereby preventing their nuclear egress and inhibiting formation of infectious virus particles. The sumoylated isoform PML-4 restricts rabies virus by inhibiting viral mRNA and protein synthesis. The cytoplasmic isoform PML-14 can restrict herpes simplex virus-1 (HHV-1) replication by sequestering the viral E3 ubiquitin-protein ligase ICP0 in the cytoplasm. Isoform PML-6 shows restriction activity towards human cytomegalovirus (HCMV) and influenza A virus strains PR8(H1N1) and ST364(H3N2). Sumoylated isoform PML-4 and isoform PML-12 show antiviral activity against encephalomyocarditis virus (EMCV) by promoting nuclear sequestration of viral polymerase (P3D-POL) within PML NBs. Isoform PML-3 exhibits antiviral activity against poliovirus by inducing apoptosis in infected cells through the recruitment and the activation of p53/TP53 in the PML-NBs. Isoform PML-3 represses human foamy virus (HFV) transcription by complexing the HFV transactivator, bel1/tas, preventing its binding to viral DNA. PML may positively regulate infectious hepatitis C viral (HCV) production and isoform PML-2 may enhance adenovirus transcription.
Ubiquitinated; mediated by RNF4, RNF111, UHRF1, UBE3A/E6AP, BCR(KLHL20) E3 ubiquitin ligase complex E3 ligase complex, SIAH1 or SIAH2 and leading to subsequent proteasomal degradation. Ubiquitination by BCR(KLHL20) E3 ubiquitin ligase complex E3 ligase complex requires CDK1/2-mediated phosphorylation at Ser-518 which in turn is recognized by prolyl-isopeptidase PIN1 and PIN1-catalyzed isomerization further potentiates PML interaction with KLHL20. 'Lys-6'-, 'Lys-11'-, 'Lys-48'- and 'Lys-63'-linked polyubiquitination by RNF4 is polysumoylation-dependent. Ubiquitination by RNF111 is polysumoylation-dependent (By similarity).
Sumoylation regulates PML's: stability in response to extracellular or intracellular stimuli, transcription directly and indirectly, through sequestration of or dissociation of the transcription factors from PML-NBs, ability to regulate apoptosis and its anti-viral activities. It is also essential for: maintaining proper PML nuclear bodies (PML-NBs) structure and normal function, recruitment of components of PML-NBs, the turnover and retention of PML in PML-NBs and the integrity of PML-NBs. Undergoes 'Lys-11'-linked sumoylation. Sumoylation on all three sites (Lys-65, Lys-160 and Lys-490) is required for nuclear body formation. Sumoylation on Lys-160 is a prerequisite for sumoylation on Lys-65. Lys-65 and Lys-160 are sumoylated by PISA1 and PIAS2. PIAS1-mediated sumoylation of PML promotes its interaction with CSNK2A1/CK2 and phosphorylation at Ser-565 which in turn triggers its ubiquitin-mediated degradation. PIAS1-mediated sumoylation of PML-RARA promotes its ubiquitin-mediated degradation. The PML-RARA fusion protein requires the coiled-coil domain for sumoylation. Sumoylation at Lys-490 by RANBP2 is essential for the proper assembly of PML-NBs. SUMO1P1/SUMO5 conjugated PML at Lys-160, Lys-380, Lys-400, Lys-490 and Lys-497, but Lys-380, Lys-400 and Lys-497 are not key acceptor lysines. SUMO1P1/SUMO5 forms polymeric chain on Lys-160 of PML by successive conjugation at 'Lys-18'; facilitating recruitment of PML-NB components, which enlarges PML. SUMO1P1/SUMO5 conjugation of PML increases SUMO2/3 conjugation, which leads to the recruitment of RNF4 and ubiquitin-dependent disintegration of PML-NBs. SUMO1P1/SUMO5 monoconjugated Lys-490. DNA damage triggers its sumoylation while some but not all viral infections can abolish sumoylation. Desumoylated by SENP1, SENP2, SENP3, SENP5 and SENP6. Arsenic induces PML and PML-RARA polysumoylation and their subsequent RNF4-dependent ubiquitination and proteasomal degradation, and is used as treatment in acute promyelocytic leukemia (APL). The nuclear isoforms (isoform PML-1, isoform PML-2, isoform PML-3, isoform PML-4, isoform PML-5 and isoform PML-6) show an increased sumoylation in response to arsenic trioxide. The cytoplasmic isoform PML-7 is not sumoylated.
Phosphorylation is a major regulatory mechanism that controls PML protein abundance and the number and size of PML nuclear bodies (PML-NBs). Phosphorylated in response to DNA damage, probably by ATR. HIPK2-mediated phosphorylation at Ser-8, Ser-36 and Ser-38 leads to increased accumulation of PML protein and its sumoylation and is required for the maximal pro-apoptotic activity of PML after DNA damage. CHEK2-mediated phosphorylation at Ser-117 is important for PML-mediated apoptosis following DNA damage. MAPK1-mediated phosphorylations at Ser-403, Ser-505, Ser-527 and Ser-530 and CDK1/2-mediated phosphorylation at Ser-518 promote PIN1-dependent PML degradation. CK2-mediated phosphorylation at Ser-565 primes PML ubiquitination via an unidentified ubiquitin ligase.
Acetylation at Lys-487 is essential for its nuclear localization. Deacetylated at Lys-487 by SIRT1 and this deacetylation promotes PML control of PER2 nuclear localization.
Nucleus. Nucleus>Nucleoplasm. Cytoplasm. Nucleus>PML body. Nucleus>Nucleolus. Endoplasmic reticulum membrane>Peripheral membrane protein>Cytoplasmic side. Early endosome membrane>Peripheral membrane protein>Cytoplasmic side.
Note: Isoform PML-1 can shuttle between the nucleus and cytoplasm. Isoform PML-2, isoform PML-3, isoform PML-4, isoform PML-5 and isoform PML-6 are nuclear isoforms whereas isoform PML-7 and isoform PML-14 lacking the nuclear localization signal are cytoplasmic isoforms. Detected in the nucleolus after DNA damage. Acetylation at Lys-487 is essential for its nuclear localization. Within the nucleus, most of PML is expressed in the diffuse nuclear fraction of the nucleoplasm and only a small fraction is found in the matrix-associated nuclear bodies (PML-NBs). The transfer of PML from the nucleoplasm to PML-NBs depends on its phosphorylation and sumoylation. The B1 box and the RING finger are also required for the localization in PML-NBs. Also found in specific membrane structures termed mitochondria-associated membranes (MAMs) which connect the endoplasmic reticulum (ER) and the mitochondria. Sequestered in the cytoplasm by interaction with rabies virus phosphoprotein.
The coiled-coil domain mediates a strong homo/multidimerization activity essential for core assembly of PML-NBs. Interacts with PKM via its coiled-coil domain (PubMed:18298799).
The B box-type zinc binding domain and the coiled-coil domain mediate its interaction with PIAS1.
Binds arsenic via the RING-type zinc finger. The RING-type zinc finger is essential for its interaction with HFV bel1/tas (PubMed:11432836).
The unique C-terminal domains of isoform PML-2 and isoform PML-5 play an important role in regulating the localization, assembly dynamics, and functions of PML-NBs.
The Sumo interaction motif (SIM) is required for efficient ubiquitination, recruitment of proteasome components within PML-NBs and PML degradation in response to arsenic trioxide.
研究領域
· Cellular Processes > Transport and catabolism > Endocytosis. (View pathway)
· Genetic Information Processing > Folding, sorting and degradation > Ubiquitin mediated proteolysis. (View pathway)
· Human Diseases > Infectious diseases: Viral > Influenza A.
· Human Diseases > Infectious diseases: Viral > Herpes simplex infection.
· Human Diseases > Cancers: Overview > Pathways in cancer. (View pathway)
· Human Diseases > Cancers: Overview > Transcriptional misregulation in cancer.
· Human Diseases > Cancers: Specific types > Acute myeloid leukemia. (View pathway)
参考文献

Application: WB Species: mouse Sample: NB4-LR1 cells
Restrictive clause
Affinity Biosciences tests all products strictly. Citations are provided as a resource for additional applications that have not been validated by Affinity Biosciences. Please choose the appropriate format for each application and consult Materials and Methods sections for additional details about the use of any product in these publications.
For Research Use Only.
Not for use in diagnostic or therapeutic procedures. Not for resale. Not for distribution without written consent. Affinity Biosciences will not be held responsible for patent infringement or other violations that may occur with the use of our products. Affinity Biosciences, Affinity Biosciences Logo and all other trademarks are the property of Affinity Biosciences LTD.